
Molecular Mechanisms Linking Obesity and Cancer
by George Bouras and Olga Tucker, MD, FRCS, FRCSI
Mr. Bouras is Senior Specialist Registrar in Surgery, and Academic Department of Surgery, University Hospital Birmingham NHS Trust, Queen Elizabeth Hospital, Birmingham, United Kingdom. Dr. Tucker is from Academic Department of Surgery, University Hospital Birmingham NHS Trust, Queen Elizabeth Hospital
Bariatric Times. 2011;8(6):16–19Financial Disclosure: The authors report no conflicts relevant to the content of this article.
ABSTRACT
The rising incidence of obesity and associated comorbidities is an increasingly important health issue in the West. Epidemiological evidence confirms an association between the presence of obesity and an increased risk of several malignancies, including endometrial, gallbladder, esophageal, renal, rectal, breast, and pancreatic cancers. Gender- and cancer-site-specific mechanisms appear to be involved, and several molecular mechanisms have been suggested. Adipocyte-secreted factors, including leptin, insulin-like growth factor 1, and inflammatory cytokines may play a role. Conversely, cancer cells may have an effect on adipocytes through secreted molecules that alter adipocyte phenotype and differentiation. This article reviews the published literature on the molecular mechanisms that appear to be involved in the epidemiological association between obesity and an increased risk of cancer. Despite our improved understanding of the mechanisms linking obesity and cancer, further research is necessary to identify clinically relevant molecular targets for cancer treatment in the context of obesity, and to raise awareness of the importance of primary prevention in this area.
INTRODUCTION
Obesity is significantly impacting public health with increased incidence of cardiovascular, respiratory, endocrine, and musculoskeletal comorbidity. Recognition of the association between obesity and cancer, particularly of the gastrointestinal tract has led to the expansion of research into the molecular mechanisms involved.[1] The possibility of a unifying mechanism responsible for carcinogenesis in individuals with obesity offers a theoretical challenge for scientists and clinicians worldwide.
EPIDEMIOLOGY
In 2006, the World Health Organization (WHO) reported 400 million adults with obesity worldwide.[2] This figure is projected to rise to over 700 million by 2015. In 2008 in the United States, 26.7 percent of adults were considered obese, an increase of 1.1 percent compared to 2007.[3] In the United Kingdom, the prevalence of obesity is similarly high; 24 and 25 percent of men and women, respectively, have obesity and 16.8 and 15.2 percent of boys and girls, respectively, aged 2 to 15 years have obesity.[4]
The association between obesity and reduction in long-term survival is well established.[5] More recently, accumulating epidemiological evidence demonstrates the association between obesity and development of malignancy. The most recent and largest meta-analysis6 revealed that an increased body mass index (BMI) of 5kg/m2 was associated with increased risk of esophageal adenocarcinoma (RR=1.52), thyroid (RR=1.33), colon (RR=1.24), and renal (RR=1.24) cancers in men. In women, a similar rise in BMI was associated with increased risk of endometrial (RR=1.59), gallbladder (RR=1.59), esophageal adenocarcinoma (RR=1.51), and renal (RR=1.34) cancers.
Weaker associations have been observed with rectal cancer, malignant melanoma, breast and pancreatic cancers. Epidemiological studies support a causal association between duration of obesity and the long latency period seen in many epithelial cancers.[7] The overall estimated population risks of incident cancers attributable to excess BMI has been estimated at 3.2 and 8.6 percent in men and women, respectively.[8]
In individuals with obesity, abdominal or visceral adiposity is associated with an increased risk of gastrointestinal tract malignancy.[9] Abdominal obesity is an independent risk factor for the development of esophageal adenocarcinoma in men.[10] Interestingly, Barrett’s esophagus, a disorder in which the lining of the esophagus is damaged by stomach acid, increases the risk of adenocarcinoma 30 to 40 times and is more closely correlated to waist circumference than raised BMI. In both colorectal and breast cancer, obesity not only increases the risk of developing cancer, but also negatively impacts the prognosis once cancer is detected.[11,12] While obesity is associated with postmenoupausal breast cancer, this is not the case in premenopausal women.[13] These findings implicate that gender- and cancer-site-specific biological mechanisms underpin the observed epidemiological associations linking obesity and malignancy. Importantly, following successful bariatric surgery, lower rates of cancer have been observed compared to age and gender matched controls.[14,15]
MOLECULAR MECHANISMS
In light of the epidemiological evidence, researchers have explored the molecular mechanisms that may be responsible for cancer promotion in individuals with obesity. Adipose tissue was previously considered an energy storage depot and mechanical barrier. Since the discovery of leptin, an adipocyte-derived pleiotropic hormone essential to appetite and weight regulation as well as reproduction, endocrine function, and immunity, in 1994,[16] over 20 adipocyte-secreted adipocytokines have been detected with evaluation of their metabolic functions, particularly insulin resistance.[17] The potential role of these adipocytokines in carcinogenesis is being explored and numerous pathways have been proposed to determine their effect.
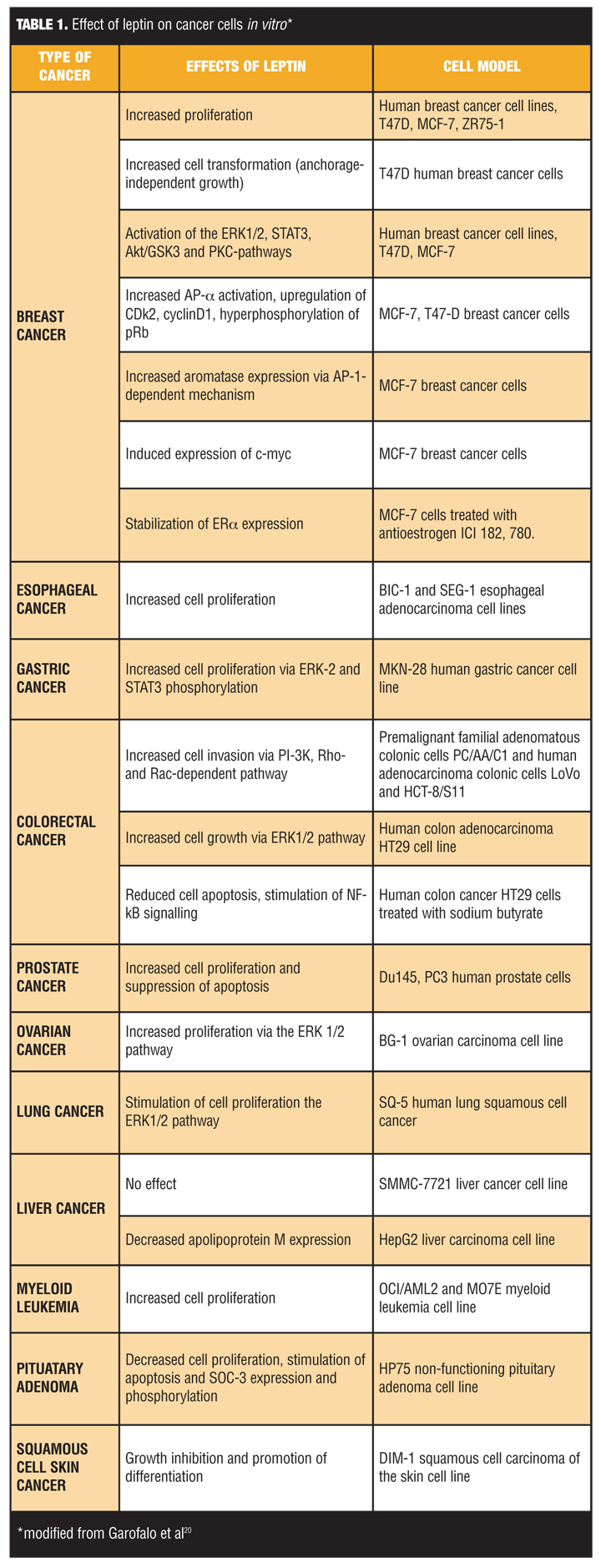
Leptin. Leptin is predominantly produced in subcutaneous white adipose tissue and is also synthesized in other types of fat and some organs, such as the liver and stomach.[18] Leptin promotes growth in many tissues and is elevated in obesity. In-vitro studies demonstrate promotion of cancer cell activity in several cancer cell lineages (Table 1).[19] Elevated serum leptin levels are observed in patients with obesity with postmenopausal breast, ovarian, and endometrial cancers.[20] In colorectal malignancy, high-serum leptin is associated with obesity, higher incidence of cancer, and poor prognosis.[21] In esophageal adenocarcinoma, obesity is associated with increased expression of leptin receptors in the primary tumor.[22] These data suggest leptin not only promotes carcinogenesis, but is also an important determinant of tumor behavior and outcome in patients with obesity.
Adiponectin. First described in 1995, adiponectin, 30 kDa complement C1-q related protein, is the most abundant gene product secreted by fat cells.[23] It is found abundantly in plasma and plays a role in glucose regulation and fatty acid catabolism. Circulating adiponectin levels are reduced in patients with obesity with promotion of insulin resistance.[24] Adiponectin has been demonstrated to have antiproliferative effects in vitro in cultured leukemia cell models.[25] Decreased serum adiponectin levels have been reported in patients with obesity with a documented malignancy.[26] In women with postmenopausal breast cancer, decreased serum adiponectin in obesity is associated with large, high-grade, primary lesions.[27] Low levels of serum adiponectin are also associated with increased risk of endometrial cancer, particularly in young women, and gastric cancer of proximal distribution.[28,29]
Preneoplastic (i.e., preceding the development of a tumor) Barrett’s esophagus has also been reported in the setting of low-serum adiponectin.[30] Laboratory-based studies confirm inhibition of cancer cell growth in esophageal and breast cancer when treated with adiponectin, an effect conveyed through increased apoptosis and inhibition of proliferation.[31,32]
Insulin-like growth factor-1. Insulin-like growth factor (IGF-1) is one of the most potent natural activators of the AKT signaling pathway, a stimulator of cell growth and proliferation, and a potent inhibitor of programmed cell death.[34] Cross-sectional studies show higher levels of IGF-1 in individuals with obesity.[35,36] Excess IGF activity through hyperinsulinemia and suppression of IGF-binding protein may promote carcinogenesis in individuals with obesity.[33] Increased serum IGF-1 levels correlate with increased risk of breast, prostate, hepatocellular, and colorectal cancer.[37–41] Tissue microarrays reveal tumor overexpression of the IGF-1 receptor as a poor prognostic marker in esophageal adenocarcinoma in patients with obesity.[42] MacDonald et al[43] recently demonstrated that a polymorphic variant of the IGF-1 receptor gene modified the risk of obesity for esophageal adenocarcinoma.[43]
Interestingly, some authors have found that weight reduction surgery significantly decreases serum IGF-1.[15,16] With increasing understanding of the functions of the IGF axis, newly discovered molecules of this cascade, including IGF-II, IGFBP2, IGFBP-3, and IGFBP-10, are being investigated with respect to obesity and carcinogensis.[44,45]
Furthermore, researchers are now exploring the effect of IGF-1 blockage on cancer cells and early clinical trials of IGF-1 receptor antagonists have shown promising results particularly in breast cancer.[46]
Inflammation and inflammatory cytokines. Obesity is associated with a systemic inflammatory response with increases in serum levels of C reactive protein, IL-6, and tumour necrosis factor (TNF)-a.[47] Obesity leads to chronic low-grade systemic inflammatory response, an environment that promotes cancer progression in many organ systems. In mice, diet-induced obesity has been demonstrated to increase systemic inflammation.[48] Co-culture of adipocytes with macrophage-conditioned media causes increased adipokine and inflammatory cytokine production by adipocytes.[49] The immune system plays an important role in the interaction between adipocytes and cancer cells, and in the induction of desmoplastic reaction, in particular. Studies investigating the effects of cancer cell culture medium on adipocytes have identified cytokines, such as TNF-a, as potent propagators of desmoplasic change.[50] Modulation of aggressiveness of prostate cancer by IL-6 from periprostatic adipose tissue has been reported.[51] The interaction between adipocytes, cancer cells, and cells of the immune system within the peri-tumoral microenvironment is not yet fully understood and requires further characterization.
Cancer cell-adipocyte interaction and pre-adipocyte differenciation. Direct in-vitro evidence of adipocyte culture medium stimulation of cancer cells has been reported in studies using breast and colon cancer lineages.[52,53] Investigators report on increased proliferation and invasion of cancer cells in response to soluble mediators secreted by adipocytes. Not only do adipocytes secrete factors that interact with cancer cells, but, conversely, the cancer cell secretome appears to influence adipocyte function.[54,55] Secreted matrix metaloprotinases and inflammatory cytokines can promote de-differentiation of mature adipocytes into preadipocytes. Preadipocytes (adipose tissue stromal cells) are particularly important as they are the most abundant mesenchymal stromal cell available for recruitment by proliferating tumor cells. They may become involved in cellular growth processes like angiogenesis.[56]
In a mouse model of breast cancer cells, migration of preadipocytes toward and into the primary cancer was observed.[57] Inflammatory mediators secreted by breast cancer cells stimulate adipocyte de-differentiation into preadipocytes and then fibroblasts leading to the induction of desmoplastic change in mammary adipose tissue.[50,54,55] This represents the first phase of connective tissue invasion by cancer cells. Interestingly, some studies have shown preadipocyte differentiation into mature adipocytes when stimulated by cancer cell culture medium.[58,59]
Physical effects of obesity. Researchers have also explored the physical effects of obesity on adipocyte function. Altered aromatase expression has been reported in adipose stromal cells of obese mice correlated with cell density in an ex-vivo model.[60] The distribution of adipocytes surrounding viscera as in abdominal adiposity plays an important role, as described previously. The effect of fat on cancer development may not only be a function of total fat volume, but also of distribution and locoregional environmental factors.
CONCLUSIONS
The obesity epidemic is continuing. In parallel, the incidence of cancer is increasing. Despite our improved understanding of the mechanisms linking obesity and cancer, further research is necessary to identify clinically relevant molecular targets for cancer treatment in the context of obesity, and to raise awareness of the importance of primary prevention in this area.
References
1. Donohoe CL, Pidgeon GP, Lysaght J, Reynolds JV. Obesity and gastrointestinal cancer. Br J Surg. 2010;97:628–642.
2. World Health Organization. Fact Sheet No.311. Obesity and Overweight. Geneva 2006, WHO.
3. CDC Vital signs: state-specific obesity prevalence among adults—United States, 2009. MMWR Morb Mortal Wkly Rep. 2010;59(30):951–955.
4. The Health and Social Care Information Centre. Statistics on obesity, physical activity and diet: England, 2010. www.ic.nhs.uk. Accessed May 19, 2011.
5. Preston SH. Deadweight? The influence of obesity on longevity. N Engl J Med. 2005;352:1135–1137.
6. Renehan AG, Tyson M, Egger M, et al. Body mass index and incidence of cancer: a systematic review and meta-analysis of prospective observational studies. Lancet. 2008; 371(9612):569–578.
7. Roberts DL, Dive C, Renehan G. Biological mechanisms linking obesity and cancer risk: new perspectives. Annu Rev Med. 2010;61:301–316.
8. Renehan AG, Soerjomataram I, Tyson M, et al. Incident cancer burden attributable to excess body mass index in 30 European countries. Int J Cancer. 2010;126(3):692–702.
9. Pischon T, Lahmann PH, Boeing H, et al. Body size and risk of colon and rectal cancer in the European Prospective Investigation Into Cancer and Nutrition (EPIC). J Natl Cancer Inst. 2006;98:920–931.
10. Corley DA, Kubo A, Zhao W. Abdominal obesity and the risk of esophageal and gastric cardia carcinomas. Cancer Epidemiol Biomarkers Prev. 2008;17(2):352–358.
11. Siegel EM, Ulrich CM, Poole EM, et al. The effects of obesity and obesity-related conditions on colorectal cancer prognosis. Cancer Control. 2010;17(1):52–57
12. Carmichael AR. Obesity as a risk factor for development and poor prognosis of breast cancer. BJOG. 2006;113(10):1160–1166.
13. Asseryanis E, Ruecklinger E, Hellan M, et al. Breast cancer size in postmenopausal women is correlated with body mass index and androgen serum levels. Gynecol Endocrinol. 2004;18:29–36.
14. Sjöström L, Gummesson A, Sjöström CD, et al. Effects of bariatric surgery on cancer incidence in obese patients in Sweden (Swedish Obese Subjects Study): a prospective, controlled intervention trial. Lancet Oncol. 2009;10(7):653–662. Epub 2009 Jun 24.
15. Adams TD, Hunt SC. Cancer and obesity: effect of bariatric surgery. World J Surg. 2009;33(10):2028–2033.
16. Zhang Y, Proenca R, Maffei M, et al. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994, 425–432.
17. Fukuhara A, Matsuda M, Nishizawa M, et al. Visfatin: a protein secreted by visceral fat that mimics the effects of insulin. Science. 2005;307:426–430.
18. Margetic S, Gazzola C, Pegg GG, Hill RA. Leptin: a review of its peripheral actions and interactions. Int J Obes Relat Metab Disord. 2002;26(11):1407–1433.
19. Garofalo C, Surmacz E. Leptin and cancer. J Cell Physiol. 2005;207:12–22.
20. Tessitore L, Vizio B, Pesola D, et al. Adipocyte expression and circulating levels of leptin increase in both gynaecological and breast cancer patients. Int J Oncol. 2004;24:1529–1535.
21. Stattin P, Lukanova A, Biessy C, et al. Obesity and colon cancer: does leptin provide a link? Int J Cancer. 2004;109:149–152.
22. Howard JM, Beddy P, Ennis D, et al. Associations between leptin and adiponectin receptor upregulation, visceral obesity and tumour stage in oesophageal and junctional adenocarcionma. Br J Surg. 2010;97(7):1020–1027.
23. Díez JJ, Iglesias P. The role of the novel adipocyte-derived hormone adiponectin in human disease. Eur J Endocrinol. 2003;148(3):293–300.
24. Hoffstedt J, Arvidsson E, Sjolin E, et al. Adipose tissue adiponectin production and adiponectin serum concentration in human obesity and insulin resistance. J Clin Endocrinol Metab. 2004;89:1391–1396.
25. Yokota T, Oritani K, Takahashi I, et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood. 2000;96:1723–1732.
26. Tian YF, Chu CH, Wu MH, et al. Anthropometric measures, plasma adiponectin and breast cancer risk. Endocr Relat Cancer. 2007;14(3):669–677.
27. Mantzoros C, Petridou E, Dessypris N, et al. Adiponectin and breast cancer risk. J Clin Endocrinol Metab. 2004;89:1102–1107.
28. Petridou E, Mantzoros C, Dessypris N, et al. Plasma adiponectin concentrations in relation to endometrial cancer: a case-control study in Greece. J Clin Endocrinol Metab. 2003;88(3):993–997.
29. Ishikawa M, Kitayama J, Kazama S, et al. Plasma adiponectin and gastric cancer. Clin Cancer Res. 2005;11:466–472.
30. Rubenstein JH, Kao JY, Madanick RD, et al. Association of adiponectin multimers with Barrett’s oesophagus. Gut. 2009;58(12):1583–1589.
31. Konturek PC, Burnat G, Rau T, et al. Effect of adiponectin and ghrelin on apoptosis of Barrett adenocarcinoma cell line. Dig Dis Sci. 2008;53(3):597–605.
32. Arditi JD, Venihaki M, Karalis KP, Chrousos GP. Antiproliferative effect of adiponectin on MCF7 breast cancer cells: a potential hormonal link between obesity and cancer. Horm Metab Res. 2007;39(1):9-13.
33. Pisani P. Hyper-insulinaemia and cancer, meta-analysis of epidemiological studies. Arch Physiol Biochem. 2008;114:63–70.
34. Nakae J, Kido Y, Accili D. Distinct and overlapping functions of insulin and IGF-1 receptors. Endocr Rev. 2001;22:818–835.
35. Gomez JM, Maravall FJ, Gomez N, et al. Interactions between serum leptin, the insulin-like growth factor-1 system, and sex, age, anthropometric and body composition variables in healthy population randomly selected. Clin Endocrinol (Oxf). 2003;58(2):213–219.
36. Gram IT, Norat T, Rinaldi S, et al. Body mass index, waist circumference and waist-hip ratio and serum levels of IGF-1 and IGFBP-3 in European Women. Int J Obes (Lond). 2006;30(11):1623–1631.
37. Chan JM, Stampfer MJ, Giovannucci E, et al. Plasma insulin-like growth factor-I and prostate cancer risk: a prospective study. Science. 1998;279:563–566.
38. Hankinson ES, Willett WC, Colditz GA, et al. Circulating concentrations of insulin-like growth factor-I and risk of breast cancer. Lancet. 1998;351:1393–1396.
39. Giovannucci E, Pollak MN, Platz EA, et al. A prospective study of plasma insulin-like growth factor-I and binding protein-3 and risk of colorectal neoplasia in women. Cancer Epidemiol Biomarkers Prev. 2000;9:345–349.
40. Mazziotti G, Sorvillo F, Morisco F, et al. Serum insulin-like growth factor I evaluation as a useful tool for predicting the risk of developing hepatocellular carcinoma in patients with hepatitis C virus-related cirrhosis. Cancer. 2002;95(12):2539–2545.
41. Imsumran A, Adachi Y, Yamamoto H, et al. Insulin-like growth factor-I receptor as a marker for prognosis and a therapeutic target in human esophageal squamous cell carcionoma. Carcionogenesis. 2007;28(5):947–956.
42. Kalinina T, Bockhorn M, Kaifi JT, et al. Insulin-like growth factor-1 receptor as a novel prognostic marker and its implications as a co-target in the treatment of human adenocarcinoma of the esophagus. Int J Cancer. 2010;127(8):1931–1940.
43. MacDonald K, Porter GA, Guernsey DL, et al. A polymorphic variant of the insulin-like growth factor type 1 receptor gene modifies risk of obesity for esophageal adenocarcinoma. Cancer Epidemiol. 2009;33(1):37–40.
44. Probst-Hensch NM, Steiner JH, Schraml P, et al. IGFBP2 and IGFBP3 protein expressions in human breast cancer: association with hormonal factors and obesity. Clin Cancer Res. 2010;16(3):1025–1032.
45. Di Martino E, Wild CP, Rotimi O, et al. IGFBP-3 and IGFBP-10 (CYR61) up-regulation during the development of Barrett’s oesophagus and associated oesophageal adenocarcionoma: potential biomarkers of disease risk. Biomarkers. 2006;11(6):547–561.
46. Tolcher AW, Sarantopoulos J, Patnaik A, et al. Phase I, pharmacokinetic, and pharmacodynamic study of AMG 479, a fully human monoclonal antibody for insulin-like growth factor receptor 1. J Clin Oncol. 2009;27(34):5800–5807.
47. Park HS, Park JY, Yu R. Relationship of obesity and visceral adiposity with serum concentrations of CRP, TNF-alpha and IL-6. Diabetes Res Clin Pract. 2005;69(1):29–35.
48. Fenton JI, Nunez NP, Yakar S, et al. Diet-induced adiposity alters the serum profile of inflammation in C57BL/6N mice as measured by antibody array. Diabetes Obes Metab. 2009;11(4):343–354.
49. Bassols J, Ortega FJ, Moreno-Navarrete JM, et al. Study of the proinflammatory role of human differentiated omental adipocytes. J Cell Biochem. 2009;107:1107–1117.
50. Meng L, Zhou J, Sasano H, et al. Tumor necrosis factor alpha and interleukin 11 secreted by malignant breast epithelial cells inhibit adipocyte differentiation by selectively down-regulating CCAAT/enhancer binding protein alpha and peroxisome proliferator-activated receptor gamma: mechanism of desmoplastic reaction. Cancer Res. 2001;61:2250–2255.
51. Finley DS, Calvert VS, Inokuchi J, et al. Periprostatic adipose tissue as a modulator of prostate cancer aggressiveness. J Urol. 2009;182(4):1621–1627.
52. Walter M, Liang S, Ghosh S, et al. Interleukin 6 secreted from adipose stromal cells promotes migration and invasion of breast cancer cells. Oncogene. 2009;28(30):2745–2755.
53. Schnäbele K, Roser S, Rechkemmer G, et al. Effects of adipocyte-secreted factors on cell cycle progression in HT29 cells. Eur J Nutr. 2009;48(3):154–161.
54. Guerrero J, Tobar N, Cáceres M, et al. Soluble factors derived from tumor mammary cell lines induce a stromal mammary adipose reversion in human and mice adipose cells. Possible role of TGF-beta1 and TNF-alpha. Breast Cancer Res Treat. 2010;119(2):497–508.
55. Andarawewa KI, Motrescu ER, Chenard MP, et al. Stromelysin-3 is a potent negative regulator of adipogenesis participating to cancer cell-adipocyte interaction/crosstalk at the tumor invasive front. Cancer Res. 2005;65(23):10862–10871.
56. Mach I, BelAiba RS, Djordjevic T, et al. Functional analysis reveal the greater potency of preadipocytes compared with adipocytes as endothelial cell activator under normoxia, hypoxia, and TNF-alpha exposure. Am J Physiol Endocrinol Metab. 2009;297:E735–E748.
57. Zhang Y, Daquinag A, Traktuev DO, et al. White adipose tissue cells are recruited by experimental tumors and promote cancer progression in mouse models. Cancer Res. 2009;69(12):5259–5266.
58. Hirano T, Morii H, Nakazawa K, et al. Enhancement of adipogenesis induction by conditioned media obtained from cancer cells. Cancer Lett. 2008;18;268(2):286–94.
59. Xu F, Gomillion C, Maxson S, Burg KJ. In vitro interaction between mouse breast cancer cells and mouse mesenchymal stem cells during adipocyte differentiation. J Tissue Eng Regen Med. 2009;3(5):338–347.
60. Ghosh S, Hu Y, Li R. Cell density is a critical determinant of aromatase expression in adipose stromal cells. J Steroid Biochem Mol Biol. 2010;118(4-5):231–236.
Category: Past Articles, Review
Subscribe
If you enjoyed this article, subscribe to receive more just like it.



{kind=link}